
Todo el contenido de iLive se revisa médicamente o se verifica para asegurar la mayor precisión posible.
Tenemos pautas de abastecimiento estrictas y solo estamos vinculados a sitios de medios acreditados, instituciones de investigación académica y, siempre que sea posible, estudios con revisión médica. Tenga en cuenta que los números entre paréntesis ([1], [2], etc.) son enlaces a estos estudios en los que se puede hacer clic.
Si considera que alguno de nuestros contenidos es incorrecto, está desactualizado o es cuestionable, selecciónelo y presione Ctrl + Intro.
Causas y patogénesis de la difteria
Médico experto del artículo.

Último revisado: 04.07.2025
La difteria es un paradigma de las enfermedades infecciosas toxigénicas. En 1883, Klebs demostró que Corynebacterium diphtheriae era el agente causal de la difteria. Un año después, Loeffler descubrió que el organismo solo podía cultivarse en la cavidad nasofaríngea y propuso que el daño a los órganos internos se debía a una toxina soluble. Para 1888, Roux y Yersin demostraron que los animales inyectados con filtrados estériles de C. diphtheriae desarrollaban una patología orgánica indistinguible de la de la difteria humana; esto demostró que una potente exotoxina era el principal factor de virulencia.
La difteria es, con mayor frecuencia, una infección de las vías respiratorias superiores que causa fiebre, dolor de garganta y malestar general. Una membrana gruesa de fibrina, de color gris verdoso, conocida como pseudomembrana, se forma a menudo en el lugar de la infección como resultado de los efectos combinados del crecimiento bacteriano, la producción de toxinas, la necrosis tisular subyacente y la respuesta inmunitaria del huésped. El reconocimiento de que el daño orgánico sistémico se debe a la acción de la toxina diftérica ha llevado al desarrollo de una terapia eficaz basada en antitoxinas para el tratamiento de la infección aguda y de una vacuna toxoide de alta eficacia.
Si bien la inmunización con toxoide ha convertido la difteria en una enfermedad poco común en zonas donde las normas de salud pública exigen la vacunación, aún se producen brotes de difteria en grupos no inmunizados e inmunodeprimidos. Por el contrario, se han producido brotes generalizados de difteria que alcanzan proporciones epidémicas en zonas donde se han suspendido los programas de inmunización activos.
Otros tipos de corinebacterias
Además de C. diphtheriae, C. ulcerans y C. pseudotuberculosis, C. pseudodiphtheriticum y C. xerosis pueden causar ocasionalmente infecciones nasofaríngeas y cutáneas. Estas dos últimas cepas se reconocen por su capacidad para producir pirazinamidasa. En veterinaria, C. renale y C. kutscheri son patógenos importantes que causan pielonefritis en el ganado bovino e infecciones latentes en ratones, respectivamente.
Causas
El agente causal de la difteria, Corynebacterium diphtheriae, es un bacilo delgado, ligeramente curvado, con engrosamientos en forma de maza en los extremos, inmóvil, no forma esporas, cápsulas ni flagelos y es grampositivo.
Además de la toxina, las corinebacterias de la difteria producen durante sus procesos vitales neuraminidasa, hialuronidasa, hemolisina, factores necrosantes y difusos, que pueden provocar necrosis y licuefacción de la sustancia principal del tejido conectivo.
Según su capacidad para formar toxinas, las corinebacterias de la difteria se dividen en toxigénicas y no toxigénicas.
La toxina diftérica es una potente exotoxina bacteriana que determina las manifestaciones clínicas generales y locales de la enfermedad. Su toxigenicidad está determinada genéticamente. Las corinebacterias no toxigénicas de la difteria no causan la enfermedad.
Según las características culturales y morfológicas, todas las corinebacterias diftéricas se dividen en tres variantes: gravis, mitis e intermedia. La gravedad de la enfermedad no depende directamente de la variante de corinebacterias diftéricas. Cada variante contiene cepas toxigénicas y no toxigénicas. Las corinebacterias toxigénicas de todas las variantes producen una toxina idéntica. [ 1 ]
Estructura, clasificación y tipos de antígenos
Corynebacterium diphtheriae es un bacilo grampositivo, inmóvil y con forma de maza. Las cepas que crecen en tejido o en cultivos in vitro más antiguos presentan puntos finos en las paredes celulares que permiten la decoloración durante la tinción de Gram y dan lugar a una reacción de Gram variable. Los cultivos más antiguos suelen contener gránulos metacromáticos (polimetafosfato) que se tiñen de púrpura azulado con azul de metileno. Los azúcares de la pared celular incluyen arabinosa, galactosa y manosa. Además, se puede aislar el éster 6,6' tóxico de trehalosa, que contiene ácidos corinemicólico y corinemicolénico en concentraciones equimolares. Se reconocen tres tipos de cultivo: mitis, intermedio y gravis.
La mayoría de las cepas requieren ácidos nicotínico y pantoténico para su crecimiento; algunas también requieren tiamina, biotina o ácido pimélico. Para una producción óptima de toxina diftérica, el medio debe suplementarse con aminoácidos y reservarse.
Ya en 1887, Loeffler describió el aislamiento de C. diphtheriae avirulentas (no toxigénicas) que eran indistinguibles de las cepas virulentas (toxigénicas) aisladas de pacientes en individuos sanos. Actualmente, se reconoce que las cepas avirulentas de C. diphtheriae pueden convertirse a un fenotipo virulento tras la infección y lisogenización por uno de varios corinebacteriófagos distintos que portan el gen estructural de la toxina diftérica, tox. La conversión lisogénica del fenotipo avirulento a virulento puede ocurrir tanto in situ como in vitro. El gen estructural de la toxina diftérica no es esencial ni para el corinebacteriófago ni para C. diphtheriae. A pesar de esta observación, no se ha observado deriva genética de la toxina diftérica.
 [
[ Patogenesia
Las vías de entrada de la infección son las mucosas de la orofaringe, la nariz y la laringe, y con menor frecuencia, las mucosas de los ojos y los genitales, así como la piel dañada, las superficies de heridas o quemaduras, la dermatitis del pañal y la herida umbilical no cicatrizada. En la vía de entrada, la corinebacteria diftérica se multiplica y secreta exotoxina.
El exudado rico en fibrinógeno se exuda y se convierte en fibrina bajo la influencia de la tromboquinasa, liberada durante la necrosis de las células epiteliales. Se forma una película fibrinosa, un signo característico de la difteria.
La portación nasofaríngea asintomática es común en las regiones endémicas de difteria. En individuos susceptibles, las cepas toxigénicas causan la enfermedad al replicarse y secretar la toxina diftérica en la nasofaringe o en lesiones cutáneas. La lesión diftérica suele estar cubierta por una pseudomembrana compuesta de fibrina, bacterias y células inflamatorias. La toxina diftérica puede escindirse proteolíticamente en dos fragmentos: el fragmento A N-terminal (dominio catalítico) y el fragmento B (dominios transmembrana y de unión al receptor). El fragmento A cataliza la ADP-ribosilación dependiente de NAD+ del factor de elongación 2, inhibiendo así la síntesis de proteínas en células eucariotas. El fragmento B se une a un receptor de la superficie celular y facilita la entrega del fragmento A al citosol.
La inmunidad protectora implica la respuesta de anticuerpos a la toxina de la difteria después de la enfermedad clínica o a la toxina de la difteria (toxina inactivada por formaldehído) después de la inmunización.
Colonización
Se sabe poco sobre los factores que median la colonización de C. diphtheriae. Sin embargo, es claro que otros factores además de la producción de toxina diftérica contribuyen a la virulencia. Estudios epidemiológicos han demostrado que un lisotipo dado puede persistir en una población durante largos períodos de tiempo. Posteriormente, puede ser reemplazado por otro lisotipo. La aparición y el dominio posterior de un nuevo lisotipo en una población está presumiblemente relacionado con su capacidad para colonizar y competir eficazmente en su segmento del nicho ecológico nasofaríngeo. Corynebacterium diphtheriae puede producir neuraminidasa, que escinde el ácido siálico de la superficie celular en sus componentes piruvato y ácido N-acetilneuramínico. El factor Cord (6,6'-di-O-micoloil-α, α'-D-trehalosa) es un componente de la superficie de C. diphtheriae, pero su papel en la colonización del huésped humano no está claro.
Producción de toxinas de difteria
El gen estructural de la toxina diftérica, tox, pertenece a una familia de corinebacteriófagos estrechamente relacionados, de los cuales el fago β es el más estudiado. La regulación de la expresión de la toxina diftérica está mediada por un represor activado por hierro, DtxR, codificado por el genoma de C. diphtheriae. La expresión de la toxina depende del estado fisiológico de C. diphtheriae. En condiciones en las que el hierro se convierte en el sustrato limitante de la tasa de crecimiento, este se disocia de DtxR, el gen toxicodendron se activa y la toxina diftérica se sintetiza y secreta al medio de cultivo a tasas máximas.
La toxina diftérica es excepcionalmente potente; para especies susceptibles (p. ej., humanos, monos, conejos, cobayas), una dosis tan pequeña como 100 a 150 ng/kg de peso corporal es letal. La toxina diftérica consiste en una sola cadena polipeptídica de 535 aminoácidos. Los análisis bioquímicos, genéticos y estructurales por rayos X muestran que la toxina consta de tres dominios estructurales/funcionales:
- ADP-ribosiltransferasa N-terminal (dominio catalítico);
- una región que facilita la entrega del dominio catalítico a través de la membrana celular (dominio transmembrana);
- dominio de unión al receptor de células eucariotas.
Tras una digestión suave con tripsina y su reducción en condiciones desnaturalizantes, la toxina diftérica puede escindirse específicamente en su asa sensible a la proteasa en dos fragmentos polipeptídicos (A y B). El fragmento A es el componente N-terminal de 21 kDa de la toxina y contiene el sitio catalítico para la ADP-ribosilación del factor de elongación 2 (EF-2).
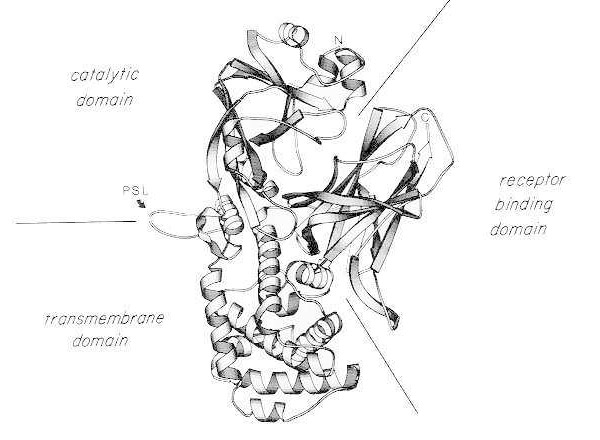
Diagrama de cinta de la estructura cristalina de rayos X de la toxina diftérica nativa monomérica. (Modificado de Bennett MJ, Choe S, Eisenberg D: Intercambio de dominios: Enredando alianzas entre proteínas. Proc Natl Acad Sci, EE. UU., 91: 3127, 1994). Se muestran las posiciones relativas de los dominios catalítico, transmembrana y de unión al receptor. La toxina intacta puede ser escindida por proteasas similares a la tripsina en Arg190, Arg192 y/o Arg193, que se encuentran en el bucle sensible a la proteasa (PSL). Tras la reducción del puente disulfuro entre Cys186 y Cys201, la toxina puede resolverse en fragmentos A y B. Se muestran los extremos amino (N) y carboxilo (C) de la toxina intacta. El diagrama de cinta se creó con el programa MOLESCRIPT.
El fragmento C-terminal, fragmento B, transporta los dominios transmembrana y de unión al receptor de la toxina.
El envenenamiento de una sola célula eucariota por la toxina de la difteria implica al menos cuatro pasos distintos:
- unión de la toxina a su receptor en la superficie celular;
- agrupamiento de receptores cargados en fosas recubiertas e internalización de la toxina a través de endocitosis mediada por receptores; después de la acidificación de la vesícula endocítica por una bomba de protones impulsada por ATP asociada a la membrana,
- inserción del dominio transmembrana en la membrana y facilitó la entrega del dominio catalítico al citosol, y
- ADP-ribosilación de EF-2, lo que resulta en una supresión irreversible de la síntesis de proteínas.
Se ha demostrado que una sola molécula del dominio catalítico entregada al citosol es suficiente para ser letal para la célula.
Diagrama esquemático de la intoxicación por difteria de una célula eucariota susceptible.
La toxina se une a su receptor de superficie celular y se internaliza a través de endocitosis mediada por receptor; tras la acidificación del endosoma, el dominio transmembrana se inserta en la membrana de la vesícula; el dominio catalítico se envía al citosol, lo que conduce a la inhibición de la síntesis de proteínas y a la muerte celular.
Epidemiología
Antes de la inmunización masiva de la población estadounidense con toxoide diftérico, la difteria era generalmente una enfermedad infantil. Un aspecto destacable de la inmunización masiva con toxoide diftérico es que, a medida que aumenta el porcentaje de la población con niveles protectores de inmunidad antitoxina (≥ 0,01 UI/ml), disminuye la frecuencia de aislamiento de cepas toxigénicas. Hoy en día, en Estados Unidos, donde la difteria clínica ha desaparecido casi por completo, el aislamiento de cepas toxigénicas de C. diphtheriae es poco frecuente. Dado que la infección subclínica ya no constituye una fuente de exposición al antígeno diftérico y, a menos que se refuerce, la inmunidad a la antitoxina disminuye, un gran porcentaje de adultos (30 a 60 %) tienen niveles de antitoxina por debajo de los niveles protectores y están en riesgo. En Estados Unidos, Europa y Europa del Este, los brotes recientes de difteria se han producido principalmente entre personas con abuso de alcohol o drogas. Dentro de este grupo, los portadores de C. diphtheriae toxigénica presentan un nivel de inmunidad antitóxica moderadamente alto. El reciente colapso de las medidas de salud pública en Rusia ha provocado que la difteria se convierta en una epidemia. A finales de 1994, se habían notificado más de 80.000 casos y más de 2.000 muertes en Rusia.
Los brotes focales de difteria casi siempre se asocian con un portador inmunitario que ha regresado de una región endémica de difteria. De hecho, brotes recientes de difteria clínica en Estados Unidos y Europa se han asociado con viajeros que regresan de Rusia y Europa del Este. Las cepas toxigénicas de C. diphtheriae se transmiten directamente de persona a persona por gotitas en el aire. Se sabe que las cepas toxigénicas pueden colonizar directamente la cavidad nasofaríngea. Además, el gen toxigénico puede propagarse indirectamente mediante la liberación de corinebacteriófagos toxigénicos y la transformación lisogénica de C. diphtheriae autóctona no toxigénica in situ. [ 5 ]
Además de la determinación del biotipo y el lisotipo de los aislados de C. diphtheriae, ahora se pueden utilizar técnicas de biología molecular para estudiar brotes de difteria. Se han empleado patrones de digestión del ADN cromosómico de C. diphtheriae mediante endonucleasas de restricción para estudiar brotes clínicos, así como secuencias de inserción de corinebacterias clonadas como sonda genética.
La prueba de Schick se ha utilizado durante muchos años para evaluar la inmunidad a la toxina diftérica, aunque ahora ha sido reemplazada en muchas áreas por la prueba serológica para anticuerpos específicos a la toxina diftérica. En la prueba de Schick, una pequeña cantidad de toxina diftérica (aproximadamente 0,8 ng en 0,2 ml) se inyecta intradérmicamente en el antebrazo (sitio de prueba) y 0,0124 μg de toxina diftérica en 0,2 ml se inyectan intradérmicamente en el sitio de control. Las lecturas se toman a las 48 y 96 horas. Las reacciones cutáneas inespecíficas suelen alcanzar su punto máximo a las 48 horas. A las 96 horas, una reacción eritematosa con alguna posible necrosis en el sitio de prueba indica inmunidad antitóxica insuficiente para neutralizar la toxina (≤ 0,03 UI/ml). La inflamación en las áreas de prueba y de control después de 48 horas indica una reacción de hipersensibilidad a la preparación de antígeno.
Formas
Corynebacterium diphtheriae infecta la nasofaringe o la piel. Las cepas toxigénicas producen una potente exotoxina que puede causar difteria. Los síntomas de la difteria incluyen faringitis, fiebre e hinchazón del cuello o la zona alrededor de la lesión cutánea. Las lesiones diftéricas están cubiertas por una pseudomembrana. La toxina se propaga por el torrente sanguíneo a órganos distantes y puede causar parálisis e insuficiencia cardíaca congestiva. [ 6 ]
Existen dos tipos de difteria clínica: nasofaríngea y cutánea. Los síntomas de la difteria faríngea varían desde faringitis leve hasta hipoxia debido a la obstrucción de las vías respiratorias por una pseudomembrana. La afectación de los ganglios linfáticos cervicales puede causar inflamación grave del cuello (difteria de cuello de toro) y el paciente puede presentar fiebre (≥ 39 °C). Las lesiones cutáneas en la difteria cutánea suelen estar cubiertas por una pseudomembrana de color marrón grisáceo. Pueden presentarse complicaciones sistémicas potencialmente mortales, principalmente pérdida de la función motora (p. ej., dificultad para tragar) e insuficiencia cardíaca congestiva, como resultado de la acción de la toxina diftérica sobre las neuronas motoras periféricas y el miocardio.
Control
El control de la difteria depende de la inmunización adecuada con toxoide diftérico: toxina diftérica inactivada por formaldehído, que permanece antigénicamente intacta. El toxoide se prepara incubando la toxina diftérica con formaldehído a 37 °C en condiciones alcalinas. La inmunización contra la difteria debe comenzar en el segundo mes de vida con una serie de tres dosis primarias a intervalos de 4 a 8 semanas, seguidas de una cuarta dosis aproximadamente 1 año después de la última primaria. El toxoide diftérico se usa ampliamente como un componente de la vacuna contra la difteria, la tos ferina y el tétanos (DPT). Los estudios epidemiológicos han demostrado que la inmunización contra la difteria tiene una efectividad de aproximadamente el 97%. Aunque la inmunización masiva contra la difteria se practica en Estados Unidos y Europa y las tasas de inmunización infantil son adecuadas, una gran proporción de la población adulta puede tener títulos de anticuerpos por debajo de los niveles protectores. Los adultos deben revacunarse con toxoide diftérico cada 10 años. De hecho, se debe administrar una inmunización de refuerzo con toxoide diftérico-tetánico a quienes viajen a áreas con altas tasas de difteria endémica (América Central y del Sur, África, Asia, Rusia y Europa del Este). En los últimos años, el uso de preparaciones de toxoides altamente purificadas para la inmunización ha minimizado las reacciones de hipersensibilidad graves ocasionales. Se debe administrar una inmunización de refuerzo con toxoide diftérico-tetánico a quienes viajen a áreas con altas tasas de difteria endémica (América Central y del Sur, África, Asia, Rusia y Europa del Este). En los últimos años, el uso de preparaciones de toxoides altamente purificadas para la inmunización ha minimizado las reacciones de hipersensibilidad graves ocasionales. Se debe administrar una inmunización de refuerzo con toxoide diftérico-tetánico a quienes viajen a áreas con altas tasas de difteria endémica (América Central y del Sur, África, Asia, Rusia y Europa del Este). En los últimos años, el uso de preparaciones de toxoides altamente purificados para la inmunización ha minimizado las reacciones de hipersensibilidad graves ocasionales.
Aunque se utilizan antibióticos (como la penicilina y la eritromicina) como parte del tratamiento de pacientes con difteria, la inmunización pasiva rápida con antitoxina diftérica es la más eficaz para reducir la mortalidad. La larga vida media de la antitoxina específica en el torrente sanguíneo es un factor importante para asegurar la neutralización eficaz de la toxina diftérica; sin embargo, para ser eficaz, la antitoxina debe reaccionar con la toxina antes de que esta pueda penetrar en la célula.
Reingeniería de la toxina de la difteria para el desarrollo de citotoxinas específicas de receptores eucariotas
La ingeniería de proteínas es un campo nuevo y en rápido desarrollo en la biología molecular; combina metodologías de ADN recombinante y síntesis de ADN en fase sólida para diseñar y construir genes quiméricos cuyos productos tienen propiedades únicas. Estudios de las relaciones estructura-función de la toxina de la difteria han demostrado claramente que esta toxina es una proteína de tres dominios: catalítico, transmembrana y receptor. Ha sido posible reemplazar genéticamente el dominio nativo de unión al receptor de la toxina de la difteria con varias hormonas polipeptídicas y citocinas (p. ej., hormona estimulante de melanocitos α [α-MSH], interleucina (IL) 2, IL-4, IL-6, IL-7, factor de crecimiento epidérmico). Las proteínas quiméricas resultantes o toxinas de fusión combinan la especificidad de unión al receptor de la citocina con los dominios transmembrana y catalítico de la toxina. En cada caso, se demostró que las toxinas de fusión envenenan selectivamente solo aquellas células que portan el receptor diana correspondiente. La primera de estas toxinas de fusión modificadas genéticamente, DAB 389IL-2, se está evaluando actualmente en ensayos clínicos en humanos para el tratamiento de linfomas refractarios y enfermedades autoinmunes en las que las células con receptores de IL-2 de alta afinidad desempeñan un papel importante en la patogénesis.[ 7 ] La administración de DAB 389 IL-2 demostró ser segura, bien tolerada y capaz de inducir una remisión duradera de la enfermedad sin efectos secundarios graves. Es probable que las toxinas de fusión basadas en la toxina de la difteria se conviertan en nuevos agentes biológicos importantes para el tratamiento de tumores o enfermedades específicas en las que se puedan dirigir receptores específicos de la superficie celular.